Microbial Variations [Genetic Manipulation in Microorganisms]
| Home | | Pharmaceutical Microbiology | | Pharmaceutical Microbiology |Chapter: Pharmaceutical Microbiology : Microbial Genetics and Variations
Importantly, the very fundamental unit of biological relatedness prevailing predominantly in various species as well as in bacteria that reproduce sexually are invariably defined by the prevelent ability of its members to copulate with one another.
MICROBIAL
VARIATIONS [GENETIC MANIPULATION IN MICROORGANISMS]
Importantly,
the very fundamental unit of biological relatedness prevailing predominantly in
various species as well as in bacteria that reproduce sexually are
invariably defined by the prevelent ability of its members to copulate with one
another. In this manner, the species do retain their ‘basic identity’ articulately
by virtue of the fact that there exist certain natural barriers that particularly check and prevent the ensuing genetic
material existing between the unrelated-organisms.
Ulti-mately, this critical identity is retained overwhelmingly via one generation to another (i.e., sustaining the so called ‘heredity’).
It has
been well established that such organisms which reproduce asexually the basic
concept of a species solely rests upon the nature’s capability to check and
prevent the exchange of the ‘genetic
material’ occurring amongst the ‘unrelated members’. One may, however,
come across the above phenomenon
quite abundantly amongst the microorganisms even though they occupy the same
kind of habitat.
Example : E.
coli and Clostridium spp. : In fact,
these two altogether divergent
organisms usually found in the ‘animal gut’, but these are quite
unrelated. Furthermore, they fail to exchange the ensuing ‘genetic information’, and thus enables the proper maintenance of
these species very much in an absolutely common environment. In fact, the entero-bacteria predominently exhibit
such vital restrictions that could be seen amongst these types of closely
related organisms.
Biologically Functional DNA Molecules : The
meticuolously designed tailor-made
biologically functional DNA molecules in the test-tube (i.e., in vitro) could be plausible and
feasible based upon the enough
concrete evidences pieced together with regard to the knowledge of the ‘nature of genetic material present in the living systems’. In other words,
one would safely conclude that the
construction of DNA might not only
replicate faithfully, but also
maintain its originality gracefully.
Chang et al. (1973) made an epoch making
discovery of constructing a miraculous biologically
functional DNA molecule in a test
tube which explicitely combined genetic information from two different sources.
Methodology : The design and construction of
such hybrid molecules were duly
accom-plished by carefully splicing
together the ‘segments’ of two
altogether different plasmids, and
subse-quently, inserting this composite
DNA plasmid strategically right into the pervailing E. coli cells. At this location, it replicated duly and thereby
succeeded in expressing the information of both parental plasmids.
By
adopting the identical procedural details the ribosomal genes of the toad Xenopus were strategically
introduced into the E. coli wherein these organisms not only replicated effectively but also expressed genuinely. Nevertheless, the RNA-DNA hybridization
technique duly detected the expression of
the inducted genes. Thus, the newly formed ‘DNA
composite molecules’ were termed as DNA
chimeras. These may be regarded as
the molecular counterparts of the
‘hybrid plant chimeras’ that can also be generated by ‘grafting’*. During the past couple of decades an enormous copius
volume of researches have been duly performed rather on a fast-track, and
eventually this new kind of work is termed as ‘plasmid engineering’ or more recent terminology ‘genetic engineering’.
Various Steps Involved in Gene Manipulation and Selection :
There are
in all four cardinal steps that are
intimately involved in accomplishing the most widely accepted and recognized
procedure of the gene manipulation and
selection, such as :
(1) Method
for cleavage and joining DNA molecules
from different sources,
(2) Search
for an appropriate ‘gene carrier’
which may replicate itself as well as the ‘foreign
DNA’ attached to it,
(3) Method
for introducing the composite DNA
molecule into a bacterial cell, and
(4) Method
for strategical selection for ‘clone of
recipient cells’ from a rather huge population.
Discovery of Ligases :
Ligases usually refer to — ‘the class of enzymes that catalyze the
joining of the ends of two chains of DNA’.
Khorana et al. (1970) first and foremost
discovered that the ligase
specifically produced by the bacteriophage T4 might occasionally capable of
catalyzing an end-to-end attachment
of an absolutely separated double
stranded DNA segment only if the
‘respectively ends’ of the two segments are able to recognize each other duly.
Even
though the above mentioned procedure happens to be not so rapid and efficient,
but it definitely paved the way for ‘intelligent
joining’ of the DNA molecules.
Salient Features :
The salient features of the genetic manipulation are as given
below :
(1) DNA terminals (ends) of
certain bacterial viruses may be joined together by the phenom-enon of ‘base-pairing’ existing between the
complementary sequences of such ‘nucleotides’
that are essentially present on the single
strand segment projecting from the ends of these molecules.
(2) Synthesis of longer segments of DNA could be
achieved by adopting the principle of link-ing together the DNA molecules by
means of the single strand projections
using wisdom, knowledge, and skill.
(3) Terminal transferase, a
relatively a recent and new enzyme, was discovered miraculously that exhibited the much desired
ability to add strategically the nucleotides
at the 3′-end of DNA. In fact, this remarkable
scientific gain of knowledge widely opened the flood-gate to-wards the
meticulous construction of a plethora of highly
specific DNA segments having critically the ‘single strand nucleotide molecules’ ; and, therefore, providing a potential avenue for joining the two pieces of DNA.
Example :
To link the DNA of animal virus SV40 with
the bacterial virus DNA :
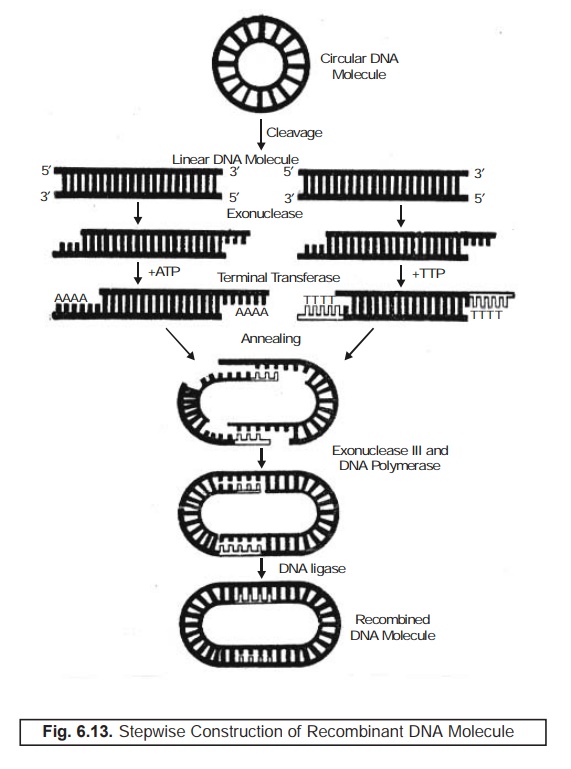
Figure :
6.13 illustrates the various steps that are involved sequentially to explain
the construction of the recombinant DNA.
(1) First
the circular DNA molecule undergoes
cleavage to yield two linear DNA molecules.
(2) Under
the influence of the enzyme ‘exonuclease’
the two fragmented linear DNA molecules give rise to terminally attached newer
elongated segments of DNA.
(3) Terminal
transferase helps these two segments
of DNA to enable them hook on further addi-tions with respective amino acids (viz., A and T).
(4) Annealing process comes
into being that specifically helps the two
loose ends of the modified linear
segments of DNA molecules to come closure in the form of a ring (not a close
ring).
(5) Presence
of exonuclease III and the DNA polymerase do help forming a circular modified DNA molecule.
(6) Finally,
the DNA ligase renders the resulting
product into a well-defined new desired ‘Recombinant
DNA Molecule’.
Generalized Procedure for Constructing Recombinant DNA Molecule and Cloning :
It has
been duly observed that the ‘biologically
active DNA’ predominantly occurs as explicitely distinct covalently-closed circles (CCC).
Nevertheless, it is first and foremost absolutely necessary to afford cleavage
of ‘circular DMA molecules' to give rise to the formation of ‘linear DNA
molecules’ having essentially free ends.
EcoRl,
and endonuclease, is observed to be an excellent enzyme most appropriate for
opening up the closed circular DNA molecules speedily and efficaciously.
Besides, it possesses the superb capability to cause an effective cleavage of
DNA at the specific sites exclusively. Subsequently, the resulting cleaved
linear DNA molecules are then duly treated with an enzyme exonuclease, which in
turn predominantly ‘chewed back' the two 5Z ends of the DNA molecules thereby
allowing the single-strand- ends projecting outwards prominently. It is
apparently followed up by the aid of the enzyme terminal transferase, whereby a
block of ‘adenines’ got hooked on to the 3Z end of one of the two DNA species
and a block of thyamines is critically attached on to the 3Z end of the other
species. Ultimately, these rw species on being mixed carefully do allow the
‘quick recognition' of the complementary blocks strategi¬cally located at the
two ends, get lined-up (i.e., alligned), and produce the much desired ‘hybrid
mol¬ecules'. At the final stage the molecules thus obtained are adequately
sealed with DNA ligases.
Figure :
6.14, depicts vividly the generalized method which is invariably adopted for
duly con-structing the recombinant DNA as well as cloning.
The
following steps summarizes the various sequential modes for obtaining the
chimeric plasmid, the transformed cell, and the daughter cells.
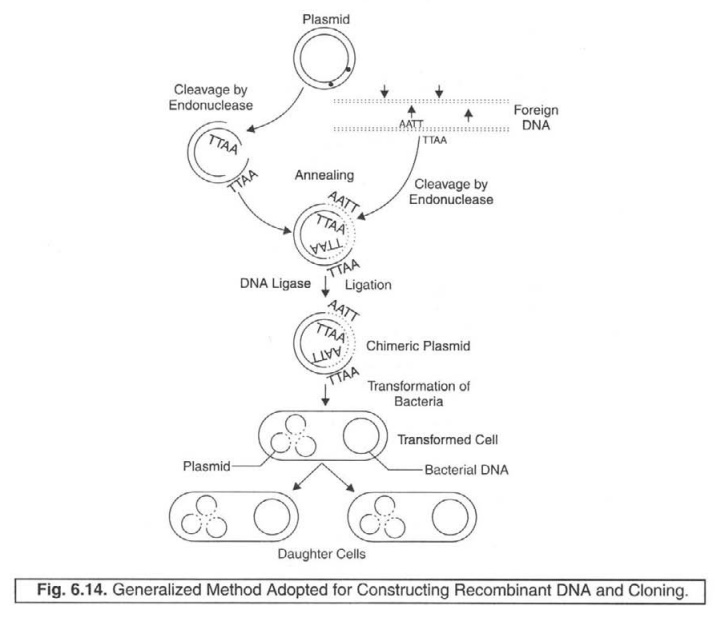
(a) Plasmid gets cleaved to corresponding
linear molecules by endonuclease ; also
accom plished by ‘foreign DNA’.
(b) Annealing process commences to obtain the
desired closed circular DNA.
(c) ‘Chimeric plasmid’ is duly
accomplished via ‘ligation’ with DNA ligase.
(d) Transformed cell is
obtained subsequently due to the
transformation of organisms.
(e) Daughter cells are
ultimately obtained from the respective
transformed cell.
Related Topics
